Схема выделения аммиака с мочой. Чем отличается мочевина от мочевой кислоты? Влияет ли избыточный вес на уровень мочевины
Аммиак – это один изконечных продуктов обмена азотсодержащих веществ. Это составляющая фракции остаточного азота сыворотки крови (наряду с мочевиной, мочевой кислотой, креатинином, индиканом). В крови концентрация аммиака невелика - 25-40 мкмоль/л. При более высоких концентрациях он оказывает токсическое действие на организм.
Аммиак токсичен, в первую очередь для ЦНС. Токсичность аммиака связана с его способностью нарушать функционирование ЦЛК, т.к. NH 3 выводит из ЦЛК–кетоглутарат:
–КГ + NH 3 + НАДН. Н + Глу + НАД + + Н 2 О.
В итогевосстановительного аминирования –кето–глутарата происходит снижение активности ЦЛК в клетках ЦНС, что, в свою очередь, угнетает активность аэробного окисления глюкозы. В результате нарушается энергопро–дукция и развивается гипоэнергетическое состояние, т.к. глюкоза – это основной источник энергии для головного мозга.
NH 3 образуется в ходе следующих процессов :
1) окислительного дезаминирования аминокислот – это основной путь продукции NH 3 ;
дезаминирования биогенных аминов;
дезаминирования пуриновых оснований (аденин, гуанин);
катаболизм пиримидиновых нуклеотидов.
В головном мозге основной источник образования NH 3 – дезаминирование АМФ до инозинмонофосфата (ИМФ):
АМФ + Н 2 О ИМФ + NH 3 .
Фермент, который катализирует эту реакцию, - аденозиндезаминаза.
Аммиак транспортируется кровью к печени и почкам для обезвреживания в составе аминокислот, среди которых основными являются глутамин, аспарагин, аланин.
Обезвреживание NH 3 происходит практически сразу после его образования, т.к. в тканях он сразу же включается в состав аминокислот, главным образом глутамина. Однако для дальнейшей детоксикации и выведения амиака существуют биохимические процессы в печени и почках, которые и являются основными путями обезвреживанияNH 3 .
Выделяют следующие механизмы обезвреживания NH 3 :
1 ) восстановительное аминирование–кетоглутарата;
2 ) образование амидов аминокислот – аспарагина и глутамина;
3 ) образование аммонийных солей в почках;
4 ) синтез мочевины.
В тканях аммиак подлежит немедленной нейтрализа–ции. Это достигается путем сочетания процессов (1 ) и (2 ).
Восстановительное аминирование –кетоглутарата :
NH 3 + –КГ + НАДН . Н + Глу + НАД + Н 2 О.
Фермент - глутаматдегидрогеназа
Для этого процесса необходимы значительные концентрации –КГ. Для того чтобы не было перерасхода–КГ и работа ЦЛК не была нарушена,–КГ пополняется за счет превращения ПВКОА–КГ.
2 )Образование амидов – это важный вспомогательный механизм обезвреживанияNH 3 в тканях путем его связывания с Глу или Асп.
Асп + АТФ + NH 3 Асн + АМФ + ФФ нн
Фермент - аспарагинсинтаза
Глу + АТФ + NH 3 Глн + АМФ + ФФ нн
Фермент - глутаминсинтаза
Этот процесс наиболее активен в ЦНС, мышцах, почках, печени (для поддержания внутренней концентрации NH 3). Главным образомглн является транспортной формой нетоксичногоNH 3 из мозга, мышц и др. тканей. Глутамин легко проникает через мембрану, т.к. при физиологических значениях рН он не имеет заряда. При физической нагрузке аланин активно транспортируетNH 3 от мышц к печени. Кроме того, большое количество аланина содержит кровь, оттекающую из кишечника. Этот аланин также направляется в печень для глюконеогенеза.
3 ) Глн и асн с током крови попадают в почки, где подвергаются гидролизу с помощью специальных ферментов – глутаминазы и аспарагиназы, которые есть и в печени:
Асн + Н 2 ОАсп +NH 3 .
Глн + Н 2 ОГлу +NH 3 .
Освободившийся в канальцах почек NH 3 нейтрализуется собразованием солей аммония, которые выводятся с мочей:
NH 3 + Н + + Сl - NH 4 Cl.
4 )Синтез мочевины - это основной путь обезвреживания аммиака. На долю мочевины приходится 80% экскретируемого азота.
Процесс образования мочевины происходит в печени и представляет собой циклический процесс, который называется «орнитиновый цикл» (цикл Кребса–Гензелайта).
В цикле принимают участие две аминокислоты, которые не входят в состав белков – орнитин и цитруллин, и две протеиногенные аминокислоты – аргинин, аспарагин.
Процесс включает пять реакций: первые две протекают в митохондриях, остальные - в цитозоле гепатоцитов. Некоторые ферменты мочевинообразования есть в мозге, эритроцитах, сердечной мышце, однако весь набор ензимов есть только в печени.
І реакция – это синтез карбамоилфосфата:
СО 2 +NH 3 + 2АТФNH 2 –CО–Ф + 2АДФ + Ф н.
Фермент - карбамоилфосфатсинтаза І (митохонд–риальный). Существует также карбамоилфосфатсинтазаІІ(в цитозоле), которая участвует в синтезе пиримидиновых нуклеотидов.
Карбамоилфосфатсинтаза І - регуляторный фермент, для которого активатором является N –ацетилглутамат .
ІІ реакция – включение карбамоилфосфата в циклический процесс. В этой реакции происходит его конденсация с орнитином), в результате чего образуется цитруллин (реакция также происходит в митохондриях).
III реакция - образование аргининосукцината. Это вторая реакция, в которой используется энергия АТФ.
IV реакция - расщепление аргининосукцината с образованием аргинина и фумарата. Последний может поступать в ЦЛК, усиливая его работу. Т.о. это анаплеротическая (пополняющая) реакция для ЦЛК.
V реакция - регенерация орнитина с образованием мочевины.
Схема синтеза мочевины
СО 2 +NH 3 + 2АТФкарбамоилфосфат + 2АДФ + Ф н
NH 2 –CО–NH 2
 (мочевина)
Орнитин
(мочевина)
Орнитин
5 2
Аргинин Цитруллин
4 3 АТФ
Фумарат АМФ
Аргининосукцинат ФФ н
Ферменты:
1 - карбамоилфосфатсинтаза;
2 - орнитинкарбамоилтрансфераза;
3 - аргининосукцинатсинтаза;
4 - аргининосукцинатлиаза;
5 - аргиназа (сильными ингибиторами фермента являются орнитин и лизин, конкурирующие с аргинином, активаторы - Са 2+ и Мn 2+).
Орнитин, который восстанавливается в ходе цикла, может запускать новый цикл мочевинообразования. По своей роли орнитин аналогичен оксалоацетату в ЦЛК. Для прохождения одного цикла необходимо 3 АТФ, которые используются в 1–й и 3–й реакциях.
Орнитиновый цикл тесно взаимосвязан с ЦЛК.
Схематически взаимосвязь можно представить так:
Орнити- СО 2
новый ЦЛК
цикл
Фумарат АТФ
Аспартат
Это «двухколесный велосипед» Кребса – ни одно колесо не способно «вращаться» без исправного функционирования второго.
Экскреция синтезированной мочевины обеспечивается почками. За сутки выделяется 20-35 г мочевины. При изменении количества белка в пище с целью поддержания азотистого равновесия скорость синтеза мочевины в организме изменяется:
белка с пищейсинтез ферментов цикласинтез мочевины,
если катаболизм белковсинтез мочевиныколичество
выводимогоазота.
Усиление катаболизма белков и, следовательно, повыше–ние экскреции мочевины наблюдаются при голодании и сахарном диабете.
При заболеваниях печени, которые сопровождаются нарушением синтеза мочевины, увеличивается концентра–ция аммиака в крови (гипераммониемия) и, как следствие, развивается печеночная кома.
Пути образования аммиака
Аммиак образуется во всех тканях, особенно в головном мозге, а обезвреживается в печени и почках.
Пути образования:
1)дезаминирование а/к-т;
2)дезаминирование пуриновых оснований;
3)распад пиримидиновых оснований;
4)дезамидирование амидов;
5)дезаминирование биогенных аминов;
6)распад гексозаминов
Дезаминирование – основной путь распада (катаболизма) а/к-т. В сутки подвергается дезаминированию ок.70 г а/к-т.
1)гидролитический
2)восстановительное дезаминирование
3)внутримолекулярное (асп,асн и гис),
4)окислительное (этот вид является основным(глу).
Процесс происходит под действием фермента – глутаматДГ, который находится в активном виде.
Пути обезвреживания аммиака .
Аммиак в малых дозах – это физиологический раздражитель, в больших – токсическое вещество. Поэтому концентрация аммиака в организме должна находиться на низком уровне – 60 мкМ/л. Гомеостаз аммиака поддерживается за счет его обезвреживания в тканях.
Пути :
1)амидирование;
2)аммонийгенез;
3)восстановительное аминирование;
4)синтез мочевины.
Аммиак, образующийся при дезамидировании в печени, обезвреживается, используясь на синтез мочевины. В почках - на образование аммонийных солей(аммонийгенез). В печени и других тканях аммиак обезвреживается в р-те реакции восстановительного аминирования. Преимущественно в печени идет процесс образования мочевины, являющийся основным путем обезвреживания аммиака.
Образование мочевины получило название орнитиновый цикл, или цитруллиновая теория. Синтез мочевины протекает в МТХ печени. Предварительно в МТХ под действием карбамоилфосфатсинтетазы I с затратой 2АТФ аммиак связывается с СО2 с образованием карбамоилфосфата:
1.В МТХ орнитинкарбамоилтрансфераза переносит карбамоильную группу карбамоилфосфата на орнитин и образуется - цитруллин:
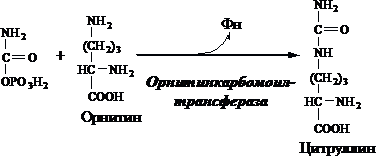
2.В цитозоле аргининосукцинатсинтетаза с затратой 1АТФ(2макроэргических связей)связывает цитруллин с аспартатом и образуется аргининосукцинат(аргининоянтарная к-та).Фермент нуждается в Mg2+.мАспартат-источник 2ого атома азота мочевины.

3.В цитозоле аргининосукцинатлиаза (аргининсукциназа) расщепляет аргининосукцинат на аргинин и фумарат (аминогруппа аспартата оказывается в аргинине).

Фумарат в ЦТК.
4.В цитозоле аргиназа гидролизует аргинин на орнитин и мочевину. У аргиназы кофакторы ионы Са2+ или Мn2+,ингибиторы-высокие концентрации орнитина и лизина.

Образующийся орнитин взаимодействует с новой молекулой карбамоилфосфата, и цикл замыкается. Мочевина в кровь->почки->выводится.
Общее уравнение синтеза мочевины:
CO2+NH3+асп+3АТФ+2Н2О→мочевина+ фумарат+2АДФ+АМФ+2Фн+ФФн.
На синтез 1 молекулы мочевины расходуются 4 макроэргических связи 3АТФ.
Ф-ии цикла:
1.превращает азота АК в мочевину, которая экскретируется и предотвращает накопление токсичных продуктов,главным образом аммиака.
2.синтез аргинина и пополнение его фонда в организме.
Нарушение цикла:
1.Гипераммониемия 1 типа - дефект карбомаилфосфат синтетазы.
2.Гипераммониемия 2типа
3.Цитруллинемия
4.Оргининсукцинатомия
5.Гиперарининемия.
Мочевинообразование - одна из важнейших дезинтоксикационных функций печени , заключающихся в преобразовании ядовитых осколков белковой молекулы в нетоксичное вещество. При дезаминировании а/к-т, нуклеотидов и др.азотистых соединений печень принимает участие в синтезе аммиака, превышение предельно допустимых концентраций которого становится высокотоксичным для организма. Дезинтоксикация аммиака происходит посредством использования его для синтеза мочевины.
Чтобы организм человека поддерживал нормальную жизнедеятельность, он выработал механизмы выведения токсических веществ. Среди них аммиак - конечный продукт метаболизма азотистых соединений, прежде всего белков. NH 3 токсичен для организма, и, как любой яд, выводится через выделительную систему. Но прежде аммиак претерпевает целый ряд последовательных реакций, который носит название орнитиновый цикл.
Типы азотистого обмена
Не для всех животных характерно выделение аммиака в окружающую среду. Альтернативными конечными веществами метаболизма азота являются и мочевина. Соответственно, называют три типа азотистого обмена, в зависимости от выделяемого вещества.
Аммониотелический тип. Конечным веществом здесь является бесцветный газ, растворимый в воде. Аммониотелия характерна для всех рыб, которые обитают в соленых водоемах.
Уреотелический тип. Животные, для которых характерна уреотелия, выделяют в окружающую среду мочевину. Примерами являются пресноводные рыбы, амфибии и млекопитающие, в том числе и человек.
Урикотелический тип. Сюда относятся те представители животного мира, у которых конечным метаболитом являются кристаллы мочевой кислоты. Это вещество как продукт азотистого обмена встречается у птиц и пресмыкающихся.
В любом из этих случаев задача конечного продукта обмена - это выведение ненужного азота из организма. Если этого не происходит, наблюдается таксация клеток и ингибирование важных реакций.
Мочевина - это амид Она образуется из аммиака, углекислого газа, азота и аминогрупп некоторых веществ во время протекания реакций орнитинового цикла. Мочевина - это продукт выделения уреотелических животных, среди которых и человек.
Мочевина является одним из способов экскреции избытка азота из организма. Образование этого вещества несет протекторную функцию, т.к. предшественник мочевины - аммиак, токсичен для клеток человека.
При переработке 100 г белка различной природы с мочой выделяется 20-25 г мочевины. Вещество синтезируется в печени, а после с током крови попадает в нефрон почки и выводится вместе с мочой.
Печень - главный орган синтеза мочевины
Во всем организме человека не найдется такой клетки, в которой будут присутствовать абсолютно все ферменты орнитинового цикла. Кроме гепатоцитов, конечно. Функция клеток печени состоит не только в синтезе и разрушении гемоглобина, но и в проведении всех реакций синтеза мочевины.
Под описание орнитинового цикла подходит тот факт, что он является единственным способом удаления азота из организма. Если на практике ингибировать синтез или действие главных ферментов, синтез мочевины остановится, а организм погибнет от переизбытка аммиака в крови.
Орнитиновый цикл. Биохимия реакций
Цикл синтеза мочевины проходит в несколько этапов. Общая схема орнитинового цикла представлена ниже (картинка), поэтому разберем каждую реакцию в отдельности. Первые две стадии протекают непосредственно в митохондриях клеток печени.
NH 3 реагирует с углекислым газом с затратой двух молекул АТФ. В результате этой энергозатратной реакции образуется карбамоилфосфат, в котором содержится макроэргическая связь. Катализирует этот процесс фермент карбамоилфосфат-синтетаза.
Карбамоилфосфат вступает в реакцию с орнитином под действием фермента орнитин-карбамоил-трансферазы. В итоге макроэргическая связь разрушается, а за счет ее энергии образуется цитруллин.
Третья и последующие стадии протекают не в митохондриях, а в цитоплазме гепатоцитов.
Идет реакция между цитруллином и аспаратом. С затратой 1 молекулы АТФ и под действием фермента аргинино-сукцинат-синтазы образуется аргинино-сукцинат.
Аргинино-сукцинат в совокупности с ферментом аргинино-сукцина-лиаза расщепляется до аргинина и фумарата.
Аргинин в присутствии воды и под действием аргиназы расщепляется до орнитина (1 реакция) и мочевины (конечный продукт). Цикл замкнулся.
Энергетика цикла синтеза мочевины
Орнитиновый цикл - энергозатратный процесс, при котором расходуются макроэргические связи молекул аденозинтрифосфата (АТФ). На протяжении всех 5 реакции в совокупности образуются 3 молекулы АДФ. К тому же энергия уходит на транспорт веществ из митохондрии в цитоплазму и наоборот. Откуда же берется АТФ?
Фумарат, который образовался в четвертой реакции, может быть использован как субстрат в цикле трикарбоновых кислот. В ходе синтеза малата из фумарата выделяется НАДФН, который дает в результате 3 молекулы АТФ.
Реакция дезаминирования глутамата также играет роль в снабжении клеток печени энергией. При этом выделяются также 3 молекулы АТФ, которые идут на синтез мочевины.
Регуляция активности орнитинового цикла
В норме каскад реакций синтеза мочевины функционирует на 60% от возможного значения. При повышенном содержании белка в пище происходит ускорение реакций, что приводит к повышению общего КПД. Метаболические нарушения орнитинового цикла наблюдаются при высоких физических нагрузках и длительном голодании, когда организм начинает расщеплять собственные белки.
Регулирование работы орнитинового цикла может происходить и на биохимическом уровне. Здесь мишенью является основной фермент карбамоилфосфат-синтетаза. Его аллостерическим активатором является N-ацетил-глутамат. При его большом содержании в организме реакции синтеза мочевины протекают нормально. При недостатке самого вещества или его предшественников, глутамата и ацетил-КоА, орнитиновый цикл теряет свою функциональную нагрузку.
Связь цикла синтеза мочевины и цикла Кребса
Реакции обоих процессов протекают в матриксе митохондрий. Это дает возможность участвовать некоторым органическим веществам в двух биохимических процессах.
СО 2 и аденозинтрифосфат, которые образуются в цикле лимонной кислоты, являются предшественниками карбамоилфосфата. АТФ также является важнейшим источником энергии.
Орнитиновый цикл, реакции которого протекают в гепатоцитах печени, - это источник фумарата, одного из важнейших субстратов в Более того, это вещество в результате нескольких поэтапных реакций дает начало аспартату, который, в свою очередь, используется в биосинтезе орнитинового цикла. Реакция с участием фумарата является источником НАДФ, с помощью которого можно фосфорилировать АДФ до АТФ.
Биологический смысл орнитинового цикла
Подавляющая часть азота поступает в организм в составе белков. В аминокислоты разрушаются, образуется аммиак как конечный продукт обменных процессов. Орнитиновый цикл - это несколько последовательных реакций, главная задача которых состоит в детоксикации NH 3 с помощью его перевода в мочевину. Мочевина, в свою очередь, поступает в нефрон почки и выводится из организма с мочой.
Кроме того, побочно орнитиновый цикл является источником аргинина - одной из незаменимых аминокислот.
Нарушения в синтезе мочевины может привести к такому заболеванию, как гипераммониемия. Эта патология характеризуется повышенной концентрацией ионов аммония NH 4 + в крови человека. Эти ионы отрицательно сказываются на жизнедеятельности организма, выключая или замедляя некоторые важные процессы. Игнорирование этого заболевания может привести к летальному исходу.
24.4.1. Биосинтез мочевины - основной путь обезвреживания аммиака. Мочевина синтезируется в орнитиновом цикле, протекающем в клетках печени. Эту последовательность реакций открыли Х.Кребс и К.Хензелейт в 1932 г. Согласно современным представлениям, цикл мочевины включает последовательность пяти реакций.
Две начальные реакции биосинтеза мочевины происходят в митохондриях клеток печени.

Последующие реакции протекают в цитоплазме клеток печени.



Общая схема орнитинового цикла представлена на рисунке 24.2:

Рисунок 24.2.
Схема орнитинового цикла и его связь с превращениями фумаровой и аспарагиновой кислот.
Цифрами обозначены ферменты, катализирующие реакции орнитинового цикла: 1 - карбамоилфосфатсинтетаза; 2 - орнитин-карбамоилтрансфераза; 3 - аргининосукцинатсинтетаза; 4 - аргининосукцинатлиаза; 5 - аргиназа.
24.4.2. Орнитиновый цикл находится в тесной взаимосвязи с циклом трикарбоновых кислот :
- пусковые реакции цикла мочевины, как и реакции ЦТК, протекают в митохондриальном матриксе;
- поступление СО2 и АТФ, необходимых для образования мочевины, обеспечивается работой ЦТК;
- в цикле мочевины образуется фумарат, который является одним из субстратов ЦТК. Фумарат гидратируется в малат, который в свою очередь окисляется в оксалоацетат. Оксалоацетат может подвергаться трансаминированию в аспартат; эта аминокислота участвует в образовании аргининосукцината.
24.4.3. Регуляция активности ферментов цикла осуществляется главным образом на уровне карбамоилфосфатсинтетазы, которая малоактивна в отсутствие своего аллостерического активатора - N-ацетил-глутамата. Концентрация последнего зависит от концентрации его предшественников (ацетил-КоА и глутамата), а также аргинина, который является аллостерическим активатором N-ацетилглутаматсинтазы:
Ацетил-КоА + Глутамат N-ацетилглутамат + КоА-SH
Концентрация ферментов орнитинового цикла зависит от содержания белка в пищевом рационе. При переходе на диету, богатую белком, в печени повышается синтез ферментов орнитинового цикла. При возвращении к сбалансированному рациону концентрация ферментов снижается. В условиях голодания, когда усиливается распад тканевых белков и использование аминокислот как энергетических субстратов, возрастает продукция аммиака, концентрация ферментов орнитинового цикла увеличивается.
24.4.4. Нарушения орнитинового цикла . Известны метаболические нарушения, обусловленные частичным блокированием каждого из 5 ферментов, катализирующих в печени реакции синтеза мочевины, а также N-ацетилглутаматсинтазы. Эти генетические дефекты, очевидно, являются частичными. Полное блокирование какой-либо из стадий цикла мочевины в печени, по-видимому, несовместимо с жизнью, потому что другого эффективного пути удаления аммиака не существует.
Общим признаком всех нарушений синтеза мочевины является повышенное содержание NH4 + в крови (гипераммониемия ). Наиболее тяжёлые клинические проявления наблюдаются при дефекте фермента карбамоилфосфатсинтетазы. Клиническими симптомами, общими для всех нарушений цикла мочевины, являются рвота, нарушение координации движений, раздражительность, сонливость и умственная отсталость. Если заболевание не диагностируется, то быстро наступает гибель. У детей старшего возраста проявлениями заболевания служат повышенная возбудимость, увеличение размеров печени и отвращение к пище с высоким содержанием белка.
Лабораторная диагностика заболеваний включает определение содержания аммиака и метаболитов орнитинового цикла в крови, моче и спинномозговой жидкости; в сложных случаях прибегают к биопсии печени.
Значительное улучшение наблюдается при ограничении белка в диете, при этом могут быть предотвращены многие нарушения мозговой деятельности. Малобелковая диета приводит к снижению содержания аммиака в крови и к улучшению клинической картины при мягких формах этих наследственных нарушений. Пищу следует принимать часто, небольшими порциями, для того чтобы избежать резкого повышения уровня аммиака в крови.
24.4.5. Клинико-диагностическое значение определения мочевины в крови и моче. В крови здорового человека содержание мочевины составляет 3,33 - 8,32 ммоль/л. За сутки с мочой выводится 20 - 35 г мочевины.
Изменения содержания мочевины в крови при заболеваниях зависят от соотношения процессов её образования в печени и выведения почками. Повышение содержания мочевины в крови (гиперазотемия) отмечается при почечной недостаточности, снижение - при недостаточности печени, при диете с низким содержанием белков.
Повышение экскреции мочевины с мочой наблюдается при употреблении пищи с высоким содержанием белков, при заболеваниях, сопровождающихся усилением катаболизма белков в тканях, при приёме некоторых лекарств (например, салицилатов). Снижение экскреции мочевины с мочой характерно для заболеваний и токсических поражений печени, заболеваний почек, сопровождающихся нарушением их фильтрационной способности.
(ОРНИТИНОВЫЙ ЦИКЛ).
Только у рыб аммиак является конечным продуктом распада и выводится из организма. У птиц и рептилий, конечный продукт катаболизма азотсодержащих соединений - мочевая кислота, а не аммиак. У млекопитающих образующийся аммиак превращается в мочевину - это полный амид угольной кислоты:
Синтез мочевины представляет собой циклический процесс, в который вступают предварительно синтезированный карбамоил-фосфат и аспартат, а образуются фумарат и мочевина.

Мочевина синтезируется из одной молекулы СО 2 , одной молекулы NH 3 и аминогруппы аспартата. Из фумарата в реакциях ЦТК вновь образуется щавелевоуксусная кислота, которая может вступать в трансаминирование с другими аминокислотами и превращаться в аспартат.
Таким образом, в орнитиновом цикле существуют два сопряженных цикла:
а) образование мочевины; б) регенерация аспартата.
В добавление к лекции по общим путям обмена аминокислот можно сказать, что еще одна, третья по счету функция трансамини рования - это перенос аминогруппы с аминокислот для синтеза мочевины без промежуточного выделения аммиака.
При синтезе мочевины расходуется в сумме 4 молекулы АТФ. Мочевина - это нетоксичное вещество, которое легко выводится из организма с мочой. Накопление мочевины в крови выше нормы происходит только при нарушениях функции почек.
Синтез мочевины происходит только в печени, а аммиак образуется в разных тканях. Значит, должен быть специальный механизм транспорта аммиака в безвредной для организма форме: это МЕХАНИЗМ ВРЕМЕННОГО ОБЕЗВРЕЖИВАНИЯ АММИАКА.
Обеспечивается ферментом глутамин-синтетазой, которая присоединяет с затратой АТФ дополнительную аминогруппу к гамма-карбоксигруппе:
 |
Аминогруппа может также присоединяться и к аспартату - к бета-карбоксигруппе.
Особенно важны реакции временного обезвреживания аммиака в нервной ткани (очень чувствительной к токсическому влиянию аммиака). Со значительной скоростью протекают они и в мышечной ткани.
Образующиеся амиды переносятся в печень, где отдают амидный азот на синтез мочевины. В почках некоторое количество аммиака из амидов может выделяться в свободном виде. Обычно такого аммиака образуется немного, но если в организме - ацидоз - сдвиг рН в кислую сторону, то этот аммиак начинает выделяться с мочой в больших количествах. Аммиак позволяет частично нейтрализовать кислоты, которые из крови при ацидозе попадают в мочу. Таким образом, глутамин и аспарагин являются транспортными формами аммиака и в составе своей молекулы переносят его из разных тканей к печени и к почкам.